
También hay una diversidad dentro de las especies: esto se llama sub-especie, raza o variedad. Sin embargo, el concepto de raza no está claro. Este término se utiliza en este artículo para referirse a un conjunto de individuos cercanos en ciertos caracteres y que tienen una historia evolutiva común en una escala más pequeña que las especies.
 |
| A• apis mellifera linnacus, B• apis mellifera koschevnicov, C •apis mellifera nigrocinta Smith, D• apis cerana Fabricius, E• apis dorsata Fabricius, F• apis florea, G• apis andreniformis |
Las diferencias entre las razas se deben a mutaciones naturales que se generalizan por la selección (natural y antropogénico), la deriva genética (probabilidad) y gracias a un flujo de genes entre razas.
No hay que confundir la raza y el ecotipo. Un ecotipo es un grupo de individuos de la misma raza y que tienen características específicas que resultan de una adaptación gradual a un entorno. Poseer un ecotipo características se heredan.
El concepto de población se utiliza muy poco; que corresponde a un conjunto de individuos de la misma especie y que tiene una ubicación común. Por tanto, una población puede corresponder a varias carreras.
En la primera parte, la metodología para la diferenciación de razas de abejas se presentará y luego a comprender la biogeografía de Apis mellifera .

Los criterios para la diferenciación de las razas de abejas
biometría
El término "biometría" significa "la medición de los vivos" y se refiere más amplio estudio cuantitativo de los seres vivos. Este artículo se basa en el trabajo de Cornuet et al. que es una síntesis de los datos biométricos en la abeja.
La realización de un examen biométrico requiere la eliminación de 30 a 40 jóvenes abejas dentro de la colmena. Deben ser mantenidas con vida y poner en una dieta de 48 horas antes de matarlas con acetato de etilo o asfixia. Las muestras se deben colocar en alcohol a 60 °.
En su estudio, Cornuet et al. utilizar métodos estadísticos planificados (análisis discriminante y la clasificación jerárquica de acuerdo con el algoritmo de Roux). Se llevaron a cabo satisfactoriamente la discriminación y clasificación de las 8 carreras de abejas y 3 híbridos interraciales (mestizos) utilizando 6 criterios morfológicos.
La coloración abdomen
La coloración abdominal se midió en el 2° Tergito abdominal (aumento x 10) y, más precisamente el ancho de tareas o la raya amarilla presente en este Tergito. La anchura obtenida debe estar entre 0,01 y 2,40 mm.

el pelo abdominal
La longitud del pelo abdominal se midió en el quinto Tergito abdominal utilizando un retículo ocular 120 divisiones (magnificación x 40).
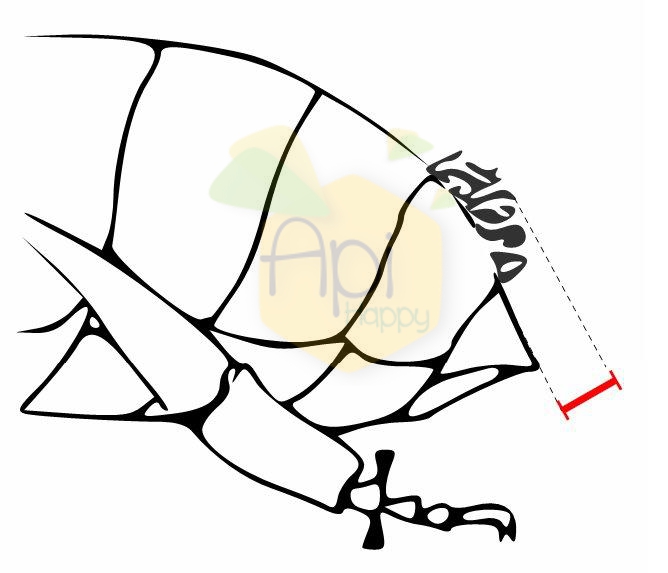
La anchura de tomento
El tomento es la banda abdominal cabelludo. Se mide en el Tergito abdominal cuarto (magnificación x 10).

La longitud de la lengua
La longitud de la lengua (probocis) se mide después de cortar y volvió la cabeza de la abeja (aumento x 10). la glosa (parte de laspiezas bucales de la abeja) se extiende utilizando unas pinzas de disección.
El índice cubital
El índice cubital se mide en las venas de las alas anteriores de un trabajador. Las nervaduras definen células. En la 3 ª celular cubital, se mide la longitud de las nervaduras A y B. El índice cubital es la relación A / B.

La transgresión discoide
La transgresión discoide se mide a las costillas de las alas anteriores de una abeja obrera. Para determinar la posición del punto discoide (A) debe trazar una línea que conecta los dos extremos de la célula radial (CR) y luego dibujar una línea a través perpendicular a la intersección de las costillas de la célula radial y cubital.
Si la línea perpendicular está en posición proximal del ala con respecto al punto A: la transgresión discoidal es negativa. Si la línea perpendicular se encuentra en una posición distal del ala con respecto al punto A: la transgresión discoidal es positiva.
Esta característica es particularmente útil para discriminar la abeja negro pero es difícil cobertura para medir puesto que es necesario determinar con precisión los extremos de la célula radial.

Resumen de los datos biométricos
Ruttner determinó en 1988 que las razas de abejas se pueden clasificar en 4 principales linajes evolutivos:
- la línea M en el oeste de Europa, desde España hasta Escandinavia (abeja negro);
- la línea C en Europa central y del norte;
- la línea A en África;
- la línea O en Turquía y el Cáucaso.
Cada una de estas líneas se ha diversificado en varias carreras. El siguiente documento presenta la distribución geográfica de cada raza y linaje. Para intermissa, la morfología, la mitocondria y los datos nucleares en realidad no se especifique la pertenencia al linaje A, o M.
La siguiente tabla resume los resultados biométricos de abejas. Los valores se han redondeado al 0,5 más cercano.
linea
|
subespecie
|
valores medios y extremos
|
Color
(mm)
|
Pilosidad
(mm)
|
Tomentum
(mm)
|
Lengua
(mm)
|
Indice cubital
|
Transgresion discoidal
|
C
(maternal)
|
Buckfast
|
medio
|
cerca
lingustica
|
cerca
lingustica
|
cerca
lingustica
|
cerca lingustica
|
cerca
lingustica
|
Positivo
|
C
|
ligustica
|
medio
|
1,75
|
0,3
|
0,85
|
6,5
|
2,3
|
Positivo
|
valor extremo
|
1,40-2,2
|
0,20-0,40
|
0,80-1,00
|
6,30-6,60
|
2,00-2,70
| |||
carnica
|
Moyenne
|
0,35
|
0,3
|
0,9
|
6,6
|
2,6
|
Positivo
| |
valor extremo
|
0,20-0,60
|
0,20-0,40
|
0,80-1,00
|
6,40-6,80
|
2,30-3,20
| |||
cecropia
|
medio
| |||||||
cypria
|
medio
|
Jaune
|
6,38
|
2.72±0.3
| ||||
O
|
anatolica
|
medio
|
2.25±0.2
| |||||
caucasica
|
medio
|
0,3
|
0,3
|
1
|
7
|
2
|
Nulo
| |
valor extremo
|
0,20-0,40
|
0,25-0,40
|
0,80-1,20
|
6,70-7,20
|
1,70-2,30
| |||
M
|
mellifera (Negra)
|
medio
|
0,25
|
0,45
|
0,75
|
6,3
|
1,75
|
Négativo
|
Valor extremo
|
0,00-0,30
|
0,40-0,52
|
0,60-0,80
|
6,00-6,50
|
1,40-2,10
| |||
A
|
intermissa
|
medio
|
0,2
|
0,2
|
0,6
|
6,4
|
2,2
| |
Valor extremo
|
0,10-0,40
|
0,15-0,35
|
0,50-0,70
|
6,30-6,60
|
2,10-2,30
| |||
lamarckii
|
medio
|
1,53
|
0,18
|
0,97
|
5,82
|
2,26
| ||
sahariensis
|
medio
|
1,53
|
0,2
|
0,53
|
5,91
|
2,71
| ||
adansonii
|
medio
|
1,52
|
0,13
|
0,37
|
5,58
|
2,2
| ||
syriava
|
medio
|
1,59
|
0,16
|
0,83
|
6,44
|
2,44
| ||
meda
|
2.5±0.2
|
Apiclass El software permite el análisis de múltiples puntos en las alas tomadas con un escáner de diapositivas (Plustek). Este software utiliza un sistema de medición semi-automática que elimina el operador efecto. Los datos han sido verificados de los análisis morfométricos y moleculares. Este sistema está disponible en línea y se requiere el uso de treinta abejas para determinar el número de miembros de una colonia para reproducirse.
Este sistema parece poco fiable (una madre negro 98% da intermissa niñas pasaron la misma foto varias veces dan resultados diferentes escáner): características de los híbridos (mestizos) no se guardan en el software.
etología
La biometría es una herramienta para identificar las razas de abejas. Hermano Adam estaba trabajando en la etología de cada carrera, es decir, su comportamiento. La siguiente tabla presenta un resumen de sus estudios.
Los siguientes cuadros son otra representación de la obra del hermano Adam. El primer lienzo compara las dos razas utilizadas principalmente por los apicultores franceses.

Podemos ver que en comparación con la abeja Buckfast, la abeja negra :
- tiene un mejor sentido de la orientación;
- construida rayos más irregulares;
- utiliza más de propóleos;
- un poco más resistente a la intemperie;
- tiene un sentido ligeramente más desarrollado del olfato;
- tiene una mayor capacidad de volar;
- tiene una durabilidad superior.
La abeja Buckfast (hermano Adán) de la abeja negra:
- tiene mejor resistencia a las enfermedades;
- tiene una mayor tendencia a pulular;
- tiene una mayor fertilidad;
- almacena la miel lejos de la camada que no bloquea la puesta;
- es más suave;
- ha ocupado la posición más alta.
estudio molecular
El estudio del genoma mitocondrial
En las abejas, la información genética está contenida en la forma de ADN en el núcleo y unos orgánulos tales como mitocondrias. Las mitocondrias se heredan única y exclusivamente de la madre. El estudio del genoma mitocondrial de un solo trabajo, por tanto, proporciona información sobre el genotipo de la reina.
El único genoma mitocondrial materno no proporcionan información sobre el genotipo de los varones de la cual los trabajadores. Este estudio proporciona sólo una determinación aproximada de la raza.
La determinación de una línea desde el genoma mitocondrial
El genoma mitocondrial permite que las líneas discrimier. Una línea corresponde a una raza o grupo de razas con una historia evolutiva común.
El genoma mitocondrial se caracteriza por las abejas regiones intergénicas CO II COI y que varían en longitud de una carrera a otra. En estas áreas, hay unidades sub P (54-67 pb, hechas de 100% adenina y timina) y subunidades Q (192-196 pb compuesto por un 93,4% y adenina . timina Estas subunidades pueden tener diferentes combinaciones: P, PQ, PQQ ... para estudiar las regiones intergénicas, hay que amplificar por PCR usando los siguientes cebadores:

Los fragmentos son entonces migran por electroforesis. La siguiente figura indica los tamaños identificados en la electroforesis en gel. Las muestras que contengan:
- subunidades son la vía P intergénica M;
- subunidades son intergénica línea Po A;
- o subunidades y P intergénicas intergénicas o subunidades Po son la línea C.

Con el fin de discriminar con precisión las subunidades intergénicas P y Po, los fragmentos de ADN deben ser digeridos por la enzima de restricción DRA1. Después de la digestión, la electroforesis se realiza usando un gel de poliacrilamida entre 5 y 10%. Haplotipos A y M se definen utilizando estos perfiles de restricción. También es posible secuenciar la porción amplificada para discriminar fragmentos P y Po.

Los datos proporcionados por el estudio del genoma mitocondrial
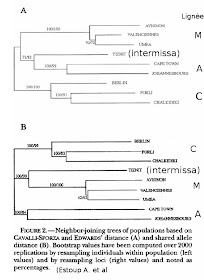
Los datos adquiridos a partir del estudio de genoma mitocondrial se utilizan en diversos métodos de clasificación y permitieron la construcción de árboles de clasificación similares. Este árbol es robusto: se hace notar en cada caso que las líneas A y C están más relacionados entre sí que con la línea M.

El estudio del genoma nuclear
El genoma nuclear se pasa a la descendencia por la madre y el padre, en el caso de l as obreras, y sólo por la madre en el caso de los zanganos ). El estudio del genoma nuclear, hacen falta al menos 50 obreras, al menos, para determinar el genotipo del macho fertilizar a la reina y, por tanto, la introgresión.
El ADN nuclear tiene ligeras variaciones entre los individuos; sólo se usan regiones altamente variables del genoma: se habla marcador. Los marcadores más utilizados en las abejas son los microsatélites. Se corresponden con zonas del genoma (loci) que consta de cientos de repeticiones en tándem unos pocos pares de bases (pb) 1-5. Por el mismo locus, hay un número variable de repeticiones de acuerdo con la subespecie (alelos).
A partir del estudio de varios loci, los árboles se hicieron por el método de fenética "vecino a participar", que cuantifica la similitud entre los individuos. Para esto, calcula un índice de similitud global y, por tanto, las distancias genéticas para cada par de individuos. Construimos el árbol por la consolidación sucesiva de los individuos más similares durante más diferentes.
Dado que el ADN es una "lengua cuaternaria" (ACTG), esta técnica plantea el problema de que dos secuencias similares pueden ser o muy cerca o muy lejos de tantas mutaciones. Para limitar el sesgo de este método, las muestras no se agrupan de acuerdo con su similitud en función, pero un algoritmo que minimiza la longitud total del eje. Con el aumento de los ordenadores potencia de cálculo, el método fenética es obsoleto y se utilizan métodos probabilísticos (sin publicación está pendiente de publicación).
Con el método de phenetic, árboles obtenidos son similares a los de los métodos mitocondriales y morfométricos excepto para la carrera intermissa que está en la línea M de acuerdo con el algoritmo utilizado.
La comprensión de la biogeografía de Apis mellifera
El trabajo del equipo Diniz Filho-JAF fue reunir datos moleculares y morfológicos para entender la biogeografía de las abejas. Su trabajo se presenta en el siguiente documento. La página central de líneas de abejas es el Oriente Medio. Notamos que :
- Una línea colonizaron África;
- línea M colonizó Europa Occidental;
- líneas C y O están más relacionados entre sí: la línea C ha colonizado los territorios al sur de los Alpes a través de Turquía, mientras que la línea O colonizaron el este de Europa a través del Cáucaso.

Los datos moleculares nos llevan a pensar que la eliminación de estas líneas tuvo lugar en el Pleistoceno. Pleistoceno, los rangos M y las líneas externas están separadas por una capa de hielo. Esto puede explicar la discrepancia surgió entre las primeras líneas y M CO. La diferencia entre C y O líneas es tarde, porque el aislamiento de las dos líneas fue más débil con el tiempo. 
Todas estas razas pueden reproducirlos. El calentamiento de Europa y el derretimiento de los casquetes polares ha llevado a una elaboración de la cerveza: las poblaciones A. mellifera tiene muchos individuos mestizos M / C de Queens importaciones de la línea C durante los últimos 40 años.
Todas estas razas pueden reproducirlos. El calentamiento de Europa y el derretimiento de los casquetes polares ha llevado a una elaboración de la cerveza: las poblaciones A. mellifera tiene muchos individuos mestizos M / C de Queens importaciones de la línea C durante los últimos 40 años.
Las razas Ibérica, y lingüstica Sicula tienen orígenes mestizos de más edad que resultan de introgresión secundaria AM ramitas (en España), MC (países del Este) y el volumen de negocios (en Francia). El mestizaje puede llegar a niveles extremos ya que la población de Portugal se compone sólo de un haplotipo A, mientras que los individuos han analizado un genoma nuclear aparentemente pura millones.
Estas diferencias pueden estar relacionados con la diversidad de comportamiento entre los sexos y razas, a la influencia del hombre y las posibles restricciones selectivas aplicadas a las moléculas estudiadas.
Conclusión sobre razas de abejas
las distintas razas de abejas no solo están determinadas por sus datos biométricos (colores, índices ...), sino también utilizando marcadores moleculares (mitocondrial y nuclear). Todo esto nos permite entender la biogeografía de la abeja.
Hay un centro de dispersión en el norte del Golfo Pérsico desde el que las abejas han colonizado Europa. La glaciación del Pleistoceno ha conducido al aislamiento de la población y el desarrollo de las razas. Después de que el hielo se derritió, las diferentes razas fueron entonces razas mixtas: la conservación y cría de razas puras, parece ser que va contra la naturaleza ahora.
Sin embargo, la preservación de las razas puras le permite disfrutar del efecto de heterosis (individuo heterocigoto)
La cría pura promueve la biodiversidad si se permite la conservación de ciertos alelos en un contexto determinado (por ejemplo, el carácter spin-off, la agresividad, la celebración de la trama). Es en este caso para crear un "banco de alelos raros" que serán necesarios en el futuro.
Bibliografía sobre razas de abejas
- Adam B (1985) - The Crossings y la apicultura mañana;
- Ruttner P. (1988) - Biogeografía y taxonomía de abejas;
- Ruttner P. (1990) The Dark Europea abeja de la miel Bibba;
- Cornuet JM et al (1975) - Discriminación y clasificación de las poblaciones de abejas de caracteres biométricos APIDOLOGIE 6 2 (1975) 145-187;
- Cornuet JM (1991) - putativos Origen y función de la zona intergénica Entre COI y CO II de Apis mellifera l. ADN mitocondrial;
- GARNERY (1992) - La historia evolutiva de la abeja de la miel Apis mellifera inferirse a partir del análisis de ADN mitocondrial;
- Estoup (1995) - la variación de microsatélites en la abeja de la miel (Apis mellifera L.) y la estructura genética de la población de prueba jerárquico de la mutación del alelo infinito y modelos paso a paso;
- Fresnaye J, (2003) - La biometría de abeja;
- Franck et al (2001) - Estudió subespecies, ubicaciones y número de COI-CO II se especifican en mitotipos Cada alquiler de muestreo. El tamaño de cada uno es mitotipo Dadas las TIC composición selon con las secuencias P, P0, P1, P2 y Q.
- GARNERY l. et al (1993) - Una prueba simple usando restringido amplificado por PCR mitochondiral ADN para estudiar la estructura genética de Apis mellifera;
- GARNERY L. et al (1998) - La diversidad genética de la abeja de la miel al oeste de Europa;
- JAF Diniz Filho et al (1999) - autocorrelación filogeográfico de la evolución fenotípica en las abejas melíferas (Apis mellifera L.);
- Franck P. (1999) - Genéticos enfoque evolutivo cuestiones asociadas con la filogeografía y la sociobiología de la abeja melífera (Apis mellifera L);
- Kekeçoglu M. (2007) Morfometría como herramienta para el estudio de la variabilidad genética de las abejas de la miel;
- Miguel et al (2007) - El flujo de genes dans le M linaje evolutivo de Apis mellifera: papel de los Pirineos, el aislamiento por distancia;
- Miguel et al (2010) - Los dos morfométricos y de microsatélites datos geométricos que apoyan consistentemente la diferenciación de los Apis mellifera M rama evolutiva y postglaciares caminos re-colonización en la Europa Occidental;
- Rortais et al (2010) - Revisión de la prueba COI-DraI CO II para la conservación de la abeja negro (Apis mellifera mellifera);
- Toullec UN K (2008) - Negro abeja, Apis mellifera mellifera, la historia y la copia de seguridad;
- Kekeçoglu M. (2009) La divergencia genética y las relaciones filogenéticas de las poblaciones de abejas de miel usando PCR-RFLP a partir del análisis de pavo de dos segmentos de ADN mitocondrial.
No hay comentarios:
Publicar un comentario