
Apis mellifera iberiensis , o la abeja española (comúnmente mal escrita como iberica ), es una subespecie de abeja occidental originaria de la Península Ibérica. También se encuentra en las Islas Baleares.
El término raza, por su parte, es fundamentalmente un concepto cultural referido a especies de animales domésticos. Aunque son insectos, las abejas melíferas (Apis mellifera, Linneo 1758) se consideran animales domésticos y, en consecuencia, podemos acudir a la zootecnia para describir diferentes razas abejas (Padilla y Flores, 2005). Desde los trabajos de Goetze (1964), las poblaciones de abejas (Apis mellifica L.) de la Península Ibérica han sido consideradas como la raza ibérica (Cornuet y Fresnaye, 1989). En el mismo sentido, Ornosa y Ortiz-Sánchez (2004), en una completa revisión de los Apoidea ibéricos, citan que la subespecie endémica de la Península Ibérica es Apis mellifera iberiensis (Engel 1999), más conocida por su antigua denominación de Apis melifera ibérica.

Finalmente, los datos nucleares también proporcionaron una evidencia convincente de que existe una barrera al flujo de genes en los Pirineos. Es decir, que la frontera geográfica entre las subespecies A. m. mellifera y A. m. iberiensis estaría en los Pirineos.Hoy en día la nomenclatura Apis mellifera iberiensis está ampliamente aceptada, tanto en la comunidad científica como en el sector apícola, para denominar a las poblaciones de abejas endémicas distribuidas a lo largo de la Península Ibérica.Las Apis mellifera iberiensis son muy vigorosas y activas a temperaturas a las que otras subespecies de abejas no salen de la colmena, soportando largos y fríos inviernos. Tienen una longitud de las alas anteriores con un promedio de 9,226 mm y 3,098 mm, mientras que el ancho de la subespecie Apis mellifera mellifera es de 9,381 mm y 3,0293 mm respectivamente.

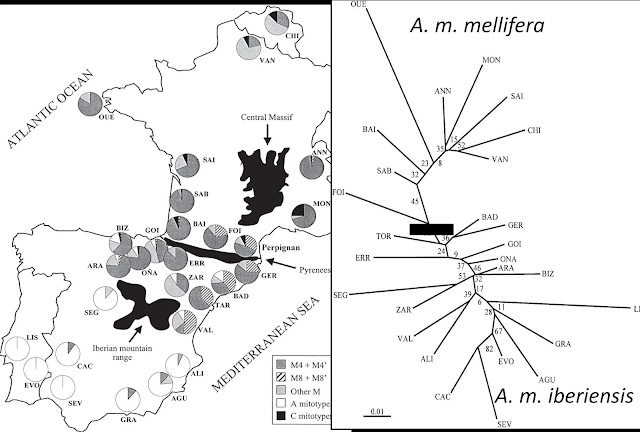

Mapas de la Península Ibérica que muestran los niveles de diversidad genética de la subespecie A. m. iberiensis. Las líneas negras continuas indican la separación entre la subespecie ibérica y las subespecies A. m. mellifera (al norte de los Pirineos) y A. m. intermissa (en el norte de África). Las líneas negras discontinuas reflejan las diferentes poblaciones de la subespecie ibérica. (A) Gradiente norte-sur de las frecuencias haplotípicas características de los linajes M (azul) y A (rojo) en base a estudios de ADN mitocondrial. La flecha diagonal indica la dirección de dicho gradiente. La flecha roja indica el sentido de la introducción artificial de poblaciones del norte de África. (B) Poblaciones de A. m. iberiensis presentes en la Península Ibérica en base a estudios de ADN nuclear.
Las actividades apícolas, especialmente el comercio de reinas, han alterado la distribución original de las subespecies de abejas melíferas (Apis mellifera) en Europa, y han dado lugar a introducciones extensivas de dos subespecies del linaje evolutivo C de la Europa oriental (A. m. ligustica y A. m. carnica) dentro del rango nativo del linaje M en la Europa Occidental (A. m. mellifera y A. m. iberiensis). Como consecuencia, en la Europa occidental se ha producido reemplazo y flujo de genes entre las poblaciones nativas y comerciales. Este fenómeno se ha producido a diferentes niveles, siendo relativamente bajo en la subespecie A. m. iberiensis en comparación a A. m. mellifera, la cual muestra un reemplazo prácticamente total en países como Alemania, Francia, Dinarmarca o Noruega.La primera descripción de esta abeja de la Península Ibérica se publicó en la revista Bee World , realizada por el hermano adam . F. Ruttner la describió en su libro Biogeografía y taxonomía de las abejas melíferas , pero al igual que varios autores anteriores a él (por ejemplo, Goetze, 1964) equiparó erróneamente a esta abeja con la subespecie propuesta por Skorikov (1929) como Apis mellifera iberica (Skorikov, sin embargo, propuso el nombre de una subespecie que se encuentra en el Cáucaso y basó el nombre en la antigua designación grecorromana para el Reino de Georgia, los íbericos caucásicos , que existía allí en la antigüedad). Por lo tanto, el nombre empleado por Ruttner fue un error, dejando a Apis mellifera iberiensis como el único nombre válido para esta subespecie de abejas melíferas. B. Adam las recogió en un viaje que realizó en 1959 a España y Portugal.
iberiensis tiene el tamaño corporal de las subespecies europeas con alas anteriores más estrechas y un abdomen más ancho. Es en su mayoría de color marrón oscuro a negro azabache. La oscuridad se acentúa por el tomento bajo y la escasa vellosidad. Las reinas son de color negro casi uniforme. Son prolíficos y de alta fertilidad controlada por las condiciones ambientales. La membrana de cierre de las células es acuosa. La raza es sensible a algunas enfermedades.

Conducta
Por lo general, no generan múltiples reinas (poliginia) en una colmena determinada en el momento del enjambre. Sus movimientos son rápidos y bastante nerviosos. Exhiben una rápida reacción defensiva, nerviosismo y una propensión a enjambrar. Hacen un uso abundante de propóleos. Una o dos abejas centinelas siempre están presentes en la entrada de la colmena. Si la colonia se ve perturbada, los centinelas emiten una alarma persistente. La colmena ataca cualquier cosa que parezca amenazante durante al menos 24 horas.
Taxonomia
El nombre que a menudo se aplica a esta raza es Amiberica , un epíteto propuesto originalmente por Skorikov en su monografía de 1929 sobre las abejas melíferas. Autores posteriores a Skorikov asumieron erróneamente que ibérica se refería a la Península Ibérica y por lo tanto rápidamente adoptaron el nombre de la raza de abejas que vivían en España y zonas limítrofes. Sin embargo, el nombre iberica se basó en una raza caucásica de abejas melíferas, el epíteto se refiere a la designación grecorromana del reino de Georgia establecido en esa región en la antigüedad. El verdadero A. m. iberica de Skorikov no tiene nada que ver con la raza de abejas del Mediterráneo occidental, y bajo las reglas de nomenclatura el nombre iberica no es válido para este linaje de abejas melíferas. El nombre corregido y válido para la raza es Apis mellifera iberiensis .
En un estudio comparativo de Amiberiensis y otras cinco subespecies de Apis mellifera , incluida Amintermissa , A. m. monticola , A. m. scutellata , A. m. adansonii y A. m. capensis [7] [8] (Smith, Palopoli, Taylor, Garnery, Cornuet, Solignac, Brown 1991) Los mapas de escisión obtenidos mediante el uso de enzimas de restricción mostraron que la abeja española contiene ADNmt similar a la intermisa y también a la mellifera . Además, Amintermissa pertenece a un grupo que se ha demostrado experimentalmente que tiene mtD.NA (ADN mitocondrial) similar, lo que incluye monticola, scuttelata, adansonii y capensis
En las poblaciones de abejas españolas, se encontró que los haplotipos de mtDNA de cepas de abejas africanas estaban presentes con frecuencia (Smith 1991, Garnery et al. 1995) (Cornuet et al 1975, 1978, 1982, 1988; Ruttner 1988; Cornuet y Fresnaye 1989; Orante-Bermejos y García-Fernández 1995; Hepburn y Radloff 1996).
Las poblaciones de abejas migratorias formaron las colonias originales de abejas en Europa occidental, aterrizando para eventualmente poblar el continente desde África a través del Estrecho de Gibraltar.
Fuente:
https://rainwaterrunoff.com/the-iberian-honey-bee/?fbclid=IwAR3i7wIAc-7_B0S8TlNb2Hc6oMdrDp30oL9fAnfOwaApryisaFWAt4LFZ80
https://www.erbel.eus/abeja-negra/
https://pubmed.ncbi.nlm.nih.gov/25930679/
Historia evolutiva de la abeja melífera ibérica (Apis mellifera iberiensis ): un enfoque del genoma completo